非小细胞肺癌 (NSCLC) 仍然是全球癌症相关死亡的主要原因,大约20-25% 的病例存在KRAS突变。KRAS-G12C突变发生在大约14% 的肺腺癌中,已成为精准医学策略的关键目标。虽然包括sotorasib和adagrasib在内的KRAS-G12C抑制剂已在临床试验中显示出希望,但其功效受到原发性和获得性耐药机制的限制。这项研究探索了将多靶点酪氨酸激酶抑制剂安洛替尼与KRAS-G12C抑制剂联合使用以克服NSCLC治疗中的这些耐药性挑战的潜力。我们的结果表明,在体外和体内,安洛替尼改善了原发性和获得性耐药环境中对KRAS-G12C抑制剂的敏感性。机械地,组合疗法抑制c-myc/ORC2信号传导,导致细胞周期停滞和细胞凋亡。这些发现表明,安洛替尼和KRAS-G12C抑制剂的组合代表了KRAS-G12C-mutant NSCLC的有希望的新治疗方法。
导言非小细胞肺癌 (NSCLC) 仍然是全球重大的健康负担,占全球癌症相关死亡的很大一部分 [
1]。NSCLC治疗中最有希望的发展之一是靶向KRAS突变,这种突变发生在20-25% 的病例中,历史上被认为是 “不可治愈的”,特别是KRAS-G12C突变 [
2,
3,
4]。KRAS-G12C突变存在于大约14% 的肺腺癌中,将其确立为精准医学方法的关键目标 [
5,
6]。KRAS-G12C抑制剂 (KRAS-G12Ci) 的开发,如sotorasib和adagrasib,已在临床试验中显示出有希望的结果,证明了先前治疗的KRAS-G12C突变NSCLC患者的适度获益,尽管总体生存率仍未改善 [
7,
8]。
KRAS-G12Ci的功效受到原发性和获得性耐药机制的限制 [
9]。在约60% 的患者中观察到的原发性耐药性可能归因于对KRAS信号缺乏依赖性 [
10]。获得性抗性通常涉及适应性非突变重编程和补偿性信号通路的反馈再激活 [
10,
11,
12]。为了应对这些挑战,已经提出了靶向多种途径的联合疗法,其中涉及KRAS-G12C抑制剂与靶向互补途径或耐药机制的药物的组合 [
12,
13]。在这种情况下,安洛替尼已成为联合治疗的有希望的候选者。
安洛替尼是一种新型的多靶点酪氨酸激酶抑制剂,已在各种癌症类型中显示出显着的抗肿瘤活性 [
14]。它主要针对血管内皮生长因子受体,成纤维细胞生长因子受体,血小板衍生生长因子受体和c-kit [
14,
15]。安罗替尼在NSCLC的临床试验中已证明有效,特别是在标准治疗后进展的患者中 [
16]。其抑制参与肿瘤生长和血管生成的多种途径的能力使其成为组合策略的有吸引力的选择。
在这项研究中,我们评估了安洛替尼与KRAS-G12Ci联合的抗肿瘤作用。我们的结果表明,这种组合抑制c-myc/起源识别复合物亚基2 (ORC2) 信号,诱导细胞周期停滞和凋亡,并且在原发性和获得性KRAS-G12Ci抗性条件下均在体外和体内发挥有效的抗肿瘤作用。这些发现确立了联合疗法作为克服KRAS-G12Ci抗性的有希望的临床策略。
方法细胞系和细胞培养在这项研究中使用了六种具有KRAS-G12C突变的NSCLC细胞系 (H2122、H2030、H358、H23、SW1573和Calu-1)。H2122和H2030购自中乔新洲生物科技 (中国上海); H358和H23购自中国科学院细胞库 (中国上海);SW1573和Calu-1均来自上海市胸科医院中心实验室。通过短串联重复序列 (STR) 分析鉴定细胞系。将细胞维持在补充有10% 胎牛血清 (GeminiBio) 、1% 青霉素-链霉素 (BasalMedia) 和1% GlutaMax (BasalMedia) 的rpmi1640培养基 (Gibco) 中。在标准培养条件下 (37 °C,5% CO2在潮湿的气氛中)。
试剂和抗体安洛替尼由正大天晴药业集团有限公司 (中国南京) 提供。Sotorasib、adagrasib和MG132购自MedChemExpress (newjersey,USA)。通过将sotorasib、adagrasib和MG132溶解在二甲基亚砜 (DMSO) 中,同时将安洛替尼溶解在无菌水中来制备储备溶液。将所有化合物储存在-20 °C下,并在使用前立即稀释至工作浓度。本研究中使用的主要抗体如下: 细胞周期蛋白B1 (Proteintech,55004-1-ap),细胞周期蛋白E1 (Proteintech,11554-1-ap),细胞周期蛋白D1 (Proteintech,60186-1-ig),CDK2 (蛋白质技术,603121-ig),GAPDH (蛋白质技术,HRP-60004),Mcl-1 (蛋白质技术,16225-1-ap),Bcl-2 (蛋白质技术,12789-1-ap),bax (克隆,A19684),Bid (abclonic,A23234),p-Erk1/2 (细胞信号传导技术,4370),Erk1/2 (细胞信号传导技术,4695),p-akt (细胞信号传导技术,4060),Akt (细胞信号传导技术,4691),β-肌动蛋白 (abclonic,AC006),c-myc (细胞信号传导技术,5605),COPS5 (abclonic,A4087),eIF4E (ab克隆,A2162),ORC2 (Abcam,A15697)。
2D培养中的细胞活力和细胞生长测定使用细胞计数试剂盒-8 (CCK-8; Dojindo实验室) 评估细胞活力。将细胞以3-5 × 10的密度接种在96孔板中3每个孔的细胞,并用指定的化合物处理72 °c。处理后,将细胞与CCK-8试剂在37 °C下孵育1和2 °C。使用酶标仪在450nm下测量吸光度。使用incucytezoom活细胞分析系统 (essenbioscience) 监测细胞生长。
协同作用分析使用多种方法量化药物联合作用。对于chou-talalay方法,使用CompuSyn软件 (1.0版) 计算组合指数 (CI) 值。CI值解释如下: CI < 1.0表示协同作用,CI = 1.0表示可加性,CI > 1.0表示拮抗作用。对于ZIP,HSA,Bliss和Loewe得分分析,根据用户教程使用Synergyfinder R软件包 (Zheng S,Wang W,Aldahdooh J 2022)。协同得分解释如下: 协同得分 < - 10表示拮抗作用;-10 < 协同得分 < 10表示可加性,协同得分 > 10表示协同作用。
集落形成试验为了评估长期细胞存活和增殖能力,以低密度接种细胞 (在六孔板中1000个细胞/孔或在96孔板中200个细胞/孔) 培养14天,每3天更换一次培养基。培养期后,吸出培养基,在室温下用4% 聚甲醛固定菌落15 ℃。固定的菌落用0.1% 结晶紫溶液染色30分钟。在用PBS温和洗涤以除去过量的污渍后,将板风干并拍照。使用ImageJ软件 (NIH,v1.54) 定量集落数目和大小。
伤口愈合试验将细胞以 ~ 80%-90% 汇合过夜接种到96孔板中,然后用IncuCyte伤口制造工具96-工具 (essenbioscience) 进行刮擦。除去分离的细胞后,补充含有所示化合物的新鲜培养基。使用IncuCyte Zoom活细胞分析系统监测和分析伤口汇合。
细胞周期和凋亡分析对于细胞周期分析,使用具有alexafluor 647 (CX004,Epizyme) 的EdU细胞增殖试剂盒,将细胞与10 μ medu在rpmi1640中孵育3小时。在用聚甲醛固定并用0.5% Triton X-100透化后,使用细胞周期染色试剂盒 (CCS012,MultiSciences) 用edu-alexafluor 647和碘化丙啶 (PI) 染色细胞。使用Attune NxT流式细胞仪 (thermofisherscientific) 进行分析。对于凋亡检测,使用膜联蛋白v-fitc/PI凋亡试剂盒 (AT101,MultiSciences),根据制造商的说明书,用膜联蛋白v-fitc和PI对细胞进行双染色。早期凋亡细胞定义为膜联蛋白V阳性/PI阴性,而晚期凋亡细胞为膜联蛋白V阳性/PI阳性。使用Flowjo软件 (flowj0llc,v10.8.1) 分析结果。
蛋白质印迹法使用补充有蛋白酶和磷酸酶抑制剂的RIPA裂解缓冲液提取总蛋白。使用二辛可宁酸测定法测定蛋白质浓度。通过10% sds-page分离等量的蛋白质并转移至PVDF膜。在室温下用TBST中的5% 脱脂乳封闭膜1 °C,然后在4 °C下与一抗孵育过夜。然后将膜与HRP缀合的第二抗体在室温下孵育1 °c。使用电化学发光成像仪观察蛋白质条带。在Image Studio软件 (li-cor) 中分析结果。
转录组测序 (rna-seq)使用TRIzol试剂 (Invitrogen) 分离三个重复的总RNA。在illuminavenovaseq6000平台上进行文库构建和配对末端测序。对原始测序数据进行质量控制,并将其映射到人类参考基因组 (GRCh38)。使用DESeq2 R软件包 (Love MI,Huber W,Anders S 2014) 进行差异表达分析,并使用基因集富集分析 (GSEA) 进行途径富集分析软件 (UC San Diego和Broad Institute,v4.1.0)。差异表达基因定义为 | log2FC | > 2并进行调整p-瓦尔 < 0.05。
RNA提取和定量PCR使用FastPure细胞/组织总RNA分离试剂盒 (RC112-01,Vazyme) 按照制造商的方案提取总RNA。使用nanodrop2000分光光度计 (thermofisherscientific) 评估RNA质量和浓度。第一链cDNA使用hiscriptivall-in-One ultratsupermix (R433-01,Vazyme) 合成。使用taqpro通用sybrqpcr主混合物 (Q712-02,Vazyme) 在LightCycler 480仪器II (Roche) 上进行实时PCR。本研究中使用的引物的5至3序列如下:
| |
|
| |
|
| |
|
| |
|
| forward GAAACCACCCCTACTCCTAATCC |
|
| 前锋TAGATCGACGTGTAGTGAATGGG |
反向TGTCTGTGGCACCAATTATTCTT |
| |
反向CTATGATACCACCCGATTGCATT |
| |
|
| |
|
| |
|
| 前进TACCAGCCCTACAGTGAAGATAC |
反向ATCAGTTTGTTGTCAGATCCTCC |
质粒和siRNA转染将细胞以40-50% 汇合接种在六孔平板中并培养过夜。对于质粒转染,根据制造商的方案将DNA与lipofectamine3000和P3000试剂 (Invitrogen) 混合。人MYC (NM_002467.6) 的质粒由GeneChem (上海,中国) 构建。对于小干扰RNA (siRNA) 转染,将寡核苷酸 (20 μ m) 与lipofectamine3000混合。转染72小时后,将细胞重新接种到六孔或96孔板中用于后续实验。通过蛋白质印迹分析验证转染效率。
5至3的ORC2 siRNA序列如下:
si-ORC2 #1: GGUUCAACAUUGUGCUUUATT
si-ORC2 #2: CCUGUUGAUAAUGGAACAUTT
细胞衍生异种移植物 (CDX) 测定和体内治疗5周龄nu/nu雌性无胸腺小鼠皮下注射1 × 107细胞悬浮在100 μ lpbs中进入侧翼区域。使用数字卡尺每3天和4天测量一次肿瘤尺寸,并使用以下公式计算肿瘤体积: 体积 = 长度 × 宽度2 × 0.5。当肿瘤达到约200 °mm时3将小鼠随机分为四个治疗组。对于h23cdx,通过胃内施用用媒介物、索托拉西布 (60 mg/kg,每天) 、安洛替尼 (2 mg/kg,每天) 或组合 (索托拉西布加安洛替尼) 处理小鼠。对于H2122SR ⑶ x,用媒介物、索托拉西布 (100 °mg/kg,每天) 、安洛替尼 (2 °mg/kg,每天) 或组合 (索托拉西布加安洛替尼) 治疗小鼠。将索托拉西布溶解在1% 吐温80、2% HPMC和97% 水中,并将安洛替尼溶解在水中。当肿瘤超过1500毫米时,对小鼠实施安乐死3或在实验终点。收获肿瘤,称重,拍照,并在聚甲醛中固定24 °c用于随后的分析。所有程序均在上海交通大学医学院动物护理和使用委员会的批准下进行。
免疫组织化学将固定的肿瘤组织石蜡包埋并切片。组织切片进行抗原修复和内源性过氧化物酶阻断,然后在4 °C与针对Ki-67、c-myc或orc2的第一抗体孵育过夜。使用ocus20显微镜载玻片扫描仪 (Grundium) 扫描切片,并使用QuPath软件 (v0.5.1) 进行分析以进行定量评估。
组织微阵列 (TMA) 分析从长沙亚翔生物技术有限公司获得了包含80例NSCLC患者样本的组织微阵列 (TMA),以及相应的临床特征和生存数据。该研究得到了公司生命科学伦理委员会 (查询代码: PZEU12C6DYBMQT) 的批准。对TMA切片进行c-myc和orc2的免疫组织化学染色。使用QuPath软件 (v0.5.1) 分析染色的切片以定量评估蛋白质表达。
统计来自体外和体内测定的定量数据表示为平均值 ± 平均值的标准误差 (SEM)。使用双尾Student's确定组之间的统计差异t-测试两组之间的比较,或单向或双向方差分析 (ANOVA),然后进行适当的事后检验以进行多重比较。相关性分析采用Pearson分析。使用graphpadprism 10软件进行所有统计分析和图形表示。对于临床数据分析,使用kaplan-meier方法生成存活曲线。使用对数秩检验评估存活的统计学差异。P-值 < 0.05被认为是统计学上显著的。
结果安洛替尼增强体外原代和获得性耐药细胞对KRAS-G12Ci的敏感性为了探索安洛替尼联合KRAS-G12Ci在KRAS-G12Ci耐药环境中的作用,我们首先研究了sotorasib对具有KRAS-G12C突变的不同NSCLC细胞系的抑制效力。细胞系对sotorasib的敏感性差异显着: H358,H2122和H2030显示出高灵敏度,而H23,Calu-1和SW1573即使在高sotorasib浓度下也能保持活力 (图
S1A)。基于这些结果,将H23、Calu-1和SW1573分类为原代抗性细胞系。
为了研究获得性抗性,我们通过将H2122和H2030细胞暴露于增加浓度的sotorasib来建立获得性抗性细胞系 (H2122SR和H2030SR) (图。
S1B)。通过细胞活力测定证实了耐药性 (图。
S1C、D)。值得注意的是,原发性和获得性耐药细胞系均保持对安洛替尼的敏感性 (图。
S1E),H2122SR和H2030SR对安洛替尼稍微更敏感 (图。
S1F)。
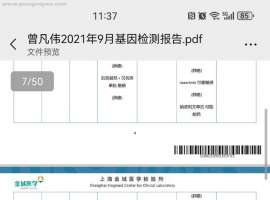
细胞活力测定和协同作用分析显示,安洛替尼剂量依赖性地增强了sotorasib在所有五种耐药细胞系 (H23,Calu-1,SW1573,H2122SR,H2030SR) 中的作用,在大多数浓度组合中观察到协同效应 (图。
1A, B,
S2A-E,和补充表
1)。当安洛替尼与另一种KRAS-G12C抑制剂adagrasib联合使用时,观察到类似的效果 (图。
S2I, J)。与单独使用任何一种药物相比,这种组合显著抑制了细胞生长 (图。
1C, D和
S2F-H)。集落形成试验表明,安洛替尼增强了索托拉西布对长期细胞存活的抑制作用 (图。
1E, F和
S2K, L)。此外,伤口愈合试验表明,该组合更有效地抑制了抗性细胞系中的细胞迁移 (图。
1G-J和
S2M-P)。这些发现共同表明,安洛替尼在体外原发性和获得性耐药环境中均增强了KRAS-G12Ci的抗肿瘤作用。
A,B安罗替尼联合索托拉西布处理的H23细胞活力和联合指数 (A) 和H2122SR (B)。结果显示为平均值 ± SEM。使用具有Tukey多重比较检验的单向ANOVA确定统计学差异。粗体:p < 0.05。n = 3每组。CI: 组合指数,受影响的Fa分数,索托拉西布,安洛安洛替尼。C,DH23的细胞生长 (C) 和H2122SR (D) 用安洛替尼 (2 μ m) 加索托拉西布 (1 μ m) 处理,使用IncuCyte监测。结果显示为平均值 ± SEM。粗体:p < 0.05。n = 3每组。组合: 安洛替尼加索托拉西布。E,FH23的集落形成测定 (E) 和H2122SR (F) 用安洛替尼 (1 μ m) 加索托拉西布 (1 μ m) 治疗14天。结果显示为平均值 ± SEM。使用具有Tukey多重比较检验的单向ANOVA确定统计学差异。粗体:p < 0.05。n = 3每组。组合: 安洛替尼加索托拉西布。G-JH23的伤口愈合测定 (G,H) 和H2122SR (我,J) 用安洛替尼 (2 μ m) 加索托拉西布 (1 μ m) 处理48 h。结果显示为平均值 ± SEM。使用具有Tukey多重比较检验的双向ANOVA确定统计学差异。粗体:p < 0.05。n = 3每组。组合: 安洛替尼加索托拉西布。
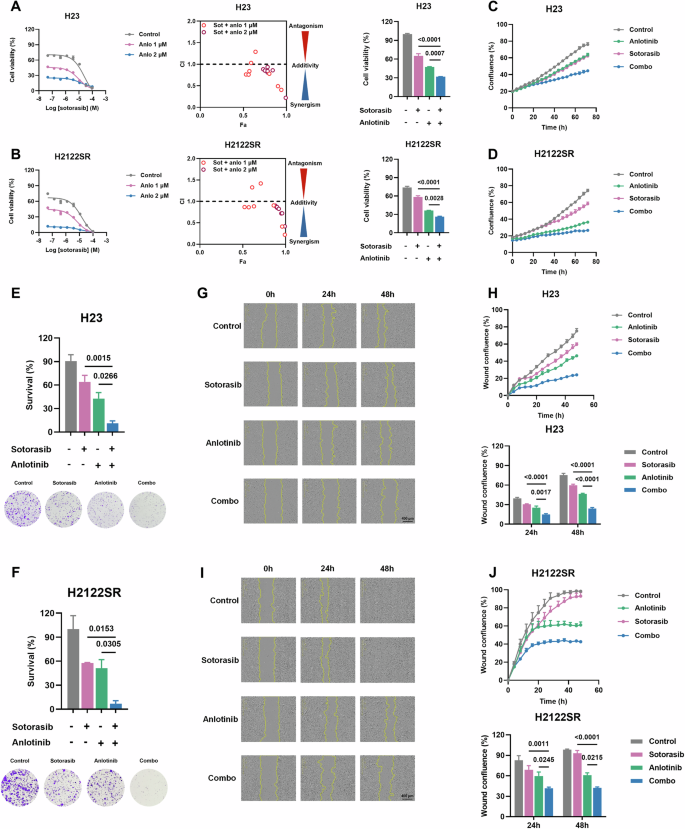
安罗替尼联合KRAS-G12Ci诱导原代及获得性耐药细胞周期阻滞和凋亡为了进一步表征联合治疗功效的潜在机制,我们检查了其对耐药NSCLC细胞系中细胞周期进程和凋亡的影响。细胞周期分析揭示了原发性和获得性抗性系之间的不同停滞模式。在原代抗性细胞系中,组合处理诱导显著的G1期停滞,而获得性抗性系显示G1和G2/M期停滞。与单药治疗相比,这两种情况都导致S期人口明显减少 (图。
2A-D和
S3A, B)。蛋白质印迹分析证实,联合治疗更有效地抑制关键细胞周期调节剂,包括细胞周期蛋白B1,细胞周期蛋白E1,细胞周期蛋白D1和CDK2,尽管抑制模式在细胞系之间有所不同 (图。
2I)。
A-DH23的EdU / PI染色测定 (A,B) 和H2122SR (C,D) 用安洛替尼 (2 μ m) 加索托拉西布 (1 μ m) 处理24.H。结果显示为平均值 ± SEM。使用具有Tukey多重比较检验的单向ANOVA确定统计学差异。粗体:p < 0.05。n = 3每组。组合: 安洛替尼加索托拉西布。E-HH23的细胞凋亡测定 (E,F) 和H2122SR (G,H) 用安洛替尼 (2 μ m) 加索托拉西布 (1 μ m) 处理24和48 h。结果显示为平均值 ± SEM。使用具有Tukey多重比较检验的双向ANOVA确定统计学差异。粗体:p < 0.05。n = 3每组。组合: 安洛替尼加索托拉西布。我用安洛替尼 (2 μ m) 处理的五种细胞系 (Calu-1,SW1573,H23,H2122SR和H2030SR) 中细胞周期蛋白B1,细胞周期蛋白E1,细胞周期蛋白D1和CDK2的表达加sotorasib (1 μ m) 24小时通过蛋白质印迹检测。索托·索托罗西布,安洛·安洛替尼。JMcl-1,Bcl-2,Bax和Bid在用安洛替尼 (2 μ m) 加索托拉西布 (1 μ m) 处理的五种细胞系 (Calu-1,SW1573,H23,H2122SR和H2030SR) 中的表达通过western印迹检测24 °h。索托·索托罗西布,安洛·安洛替尼。
凋亡的时程分析表明,与单一药物相比,联合治疗诱导的凋亡率显着提高,并且效果随时间而增加 (图
2E-H和
S3C, D)。从机制上讲,增强的凋亡反应与抑制抗凋亡蛋白 (Mcl-1和Bcl-2) 相关,而不是与促凋亡蛋白 (Bax和Bid) 的变化相关 (图。
2J)。这些发现表明,安洛替尼与KRAS-G12Ci组合通过双重机制有效地克服了原发性和获得性耐药: 诱导细胞周期停滞,随后S期减少和促进稳健的凋亡反应。
为了阐明增强的抗肿瘤作用的潜在机制,我们首先研究了抗性细胞系中经典RAS信号传导的变化。虽然索托拉西布保持了显著抑制p-erk的能力,但它对p-akt的影响很小,安洛替尼对RAS信号没有产生额外的影响 (图。
S4A)。
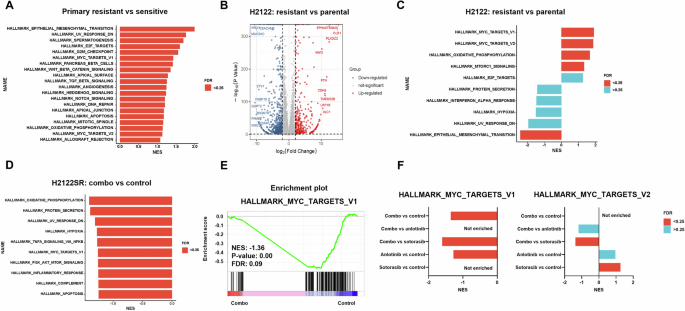
使用CCLE数据库对初级敏感细胞系和耐药细胞系进行转录组分析 [
17] 揭示了耐药细胞中多种富集的恶性肿瘤相关途径,包括EMT,E2F靶标,G2M检查点和MYC靶标 (图。
3A和
S4B)。同样,adagrasib耐药SW837 (KRAS-G12C突变型结直肠腺癌细胞系) CDX模型的GSEA [
18] 还表明,与未治疗的肿瘤相比,MYC靶点是最丰富的途径之一 (图。
S4C)。
A与敏感细胞系 (H358,H2122,H2030) 相比,原代抗性细胞系 (Calu-1,SW1573,H23) 的富集标志途径。绘制归一化富集分数 (NES)。错误发现率 (FDR) < 0.25用红色标记。数据来自CCLE数据集。B火山图显示H2122SR与H2122亲本细胞系相比的转录差异。具有对数的基因2Fc > 2并调整p值 < 0.05用红色标记。具有对数的基因2Fc < − 2并调整p值 < 0.05用蓝色标记。C与H2122亲本细胞系相比,H2122SR的富集标志途径。绘制归一化富集分数 (NES)。FDR < 0.25用红色标记。FDR > 0.25用蓝色标记。D与载体相比,用安洛替尼 (2 μ m) 加索托拉西布 (1 μ m) 处理24小时的H2122SR的富集标志途径。绘制归一化富集分数 (NES)。FDR < 0.25用红色标记。组合: 安洛替尼加索托拉西布。E与载体相比,用安洛替尼 (2 μ m) 加索托拉西布 (1 μ m) 处理24小时的H2122SR的MYC靶V1途径的GSEA富集图。FBarplot比较不同治疗组富集的MYC靶标V1和MYC靶标V2途径的NSE和FDR: DMSO,安洛替尼 (2 μ m),索托拉西布 (1 μ m),和组合 (安洛替尼加索托拉西布) 24 °h。FDR < 0.25用红色标记。FDR > 0.25用蓝色标记。组合: 安洛替尼加索托拉西布。
H2122与其抗性对应物H2122SR之间的rna-seq比较鉴定出902个上调基因和1601个下调基因 (图。
3B)。GSEA分析强调MYC靶标V1、MYC靶标V2和氧化磷酸化是H2122SR中前三个显著富集的途径 (图。
3C和
S4D)。
用sotorasib,anlotinib或它们的组合处理24小时后,H2122SR细胞的转录组分析揭示了组合处理的细胞中各种途径的富集,包括通过nf-κb的tnf-α 信号传导,MYC靶向V1和PI3K-Akt-mTOR信号 (图。
3D, E和
S4E)。
与单一药物治疗相比,联合治疗进一步下调了MYC靶通路 (图。
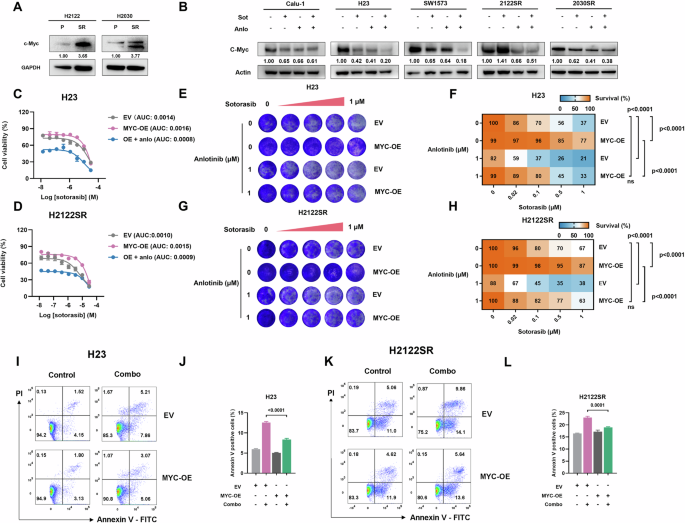
3F)。qPCR和蛋白质印迹分析证实,与亲本系相比,H2122SR和H2030SR中的MYC表达在mRNA和蛋白质水平上均显着升高 (图
4A和
S4F),而联合治疗有效抑制了所有耐药细胞系中MYC的mRNA和蛋白质水平 (图。
4B和
S4G)。值得注意的是,MYC过表达降低了耐药细胞对sotorasib的敏感性; 然而,将安洛替尼与sotorasib联合使用有效地逆转了MYC过表达对细胞活力的影响 (图。
4C, D) 和长期生存率 (图。
4E-H)。MYC过表达部分拯救了细胞免受联合治疗对细胞活力的抑制作用 (图。
S4H),长期生存 (图。
4E-H) 和细胞凋亡 (图。
4I-L)。
A,B与亲本细胞系相比,H2122SR和H2030SR中c-myc的表达 (A),并在五个细胞系 (Calu-1,SW1573,H23,H2122SR和H2030SR) 中用安洛替尼 (2 μ m) 加索托拉西布 (1 μ m) 处理24 h (B) 通过蛋白质印迹法检测。索托·索托罗西布,安洛·安洛替尼。C,DH23的细胞活力测定 (C) 和H2122SR (D) 在转染空载体 (EV) 或MYC过表达 (OE) 质粒后,单独或与安洛替尼联合使用sotorasib处理。结果显示为平均值 ± SEM。索托·索托罗西布,安洛·安洛替尼。E-H菌落形成测定显示单独或组合处理的H23的sotorasib的长期生长 (E,F) 和H2122SR (G,H) 在EV或MYC过表达后。使用具有Tukey多重比较检验的双向ANOVA确定统计学差异。粗体:p < 0.05。n = 3每组。I-LH23的细胞凋亡测定 (E,F) 和H2122SR (G,H) 在EV或MYC过表达后,用安洛替尼 (2 μ m) 加索托拉西布 (1 μ m) 处理24 h。结果显示为平均值 ± SEM。使用具有Tukey多重比较检验的单向ANOVA确定统计学差异。粗体:p < 0.05。n = 3每组。
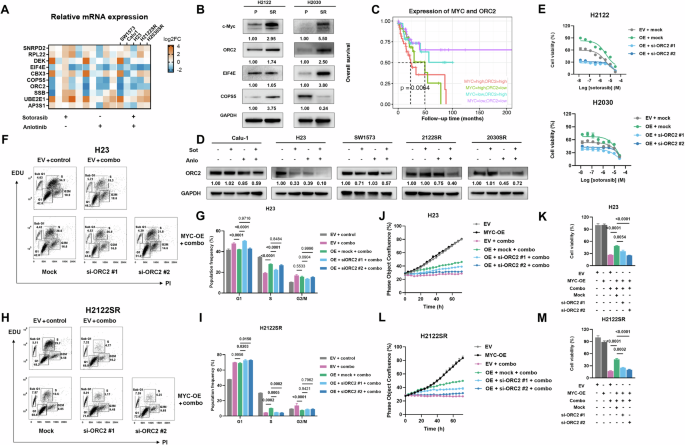
C-myc/ORC2轴是安洛替尼和KRAS-G12Ci联合治疗反应的关键信号通路为了鉴定c-myc在介导联合治疗反应中的关键下游效应子,我们分析了Myc靶标途径中前10个最下调的基因。耐药细胞系的qPCR验证显示,EIF4E、COPS5和ORC2在联合治疗24小时后一致下调 (图。
5A)。而MYC过表达在mRNA水平上调了所有三个靶点 (图。
S4I),与亲本品系相比,H2122SR和H2030SR中只有ORC2表达升高 (图。
5B)。蛋白酶体抑制实验表明,MG132稳定了c-myc蛋白的表达。此外,单独或与sotorasib联合使用的安洛替尼的抑制作用被部分逆转,表明这些作用至少部分被介导,蛋白质稳定性也降低 (图。
S4J)。
A五种细胞系 (Calu-1,SW1573,H23,H2122SR和H2030SR) 中SNRPD2,RPL22,DEK,EIF4E,COPS5,ORC2,SSB,UBE2E1,AP3S1的mRNA表达热图通过qPCR检测。n = 3每组。B通过蛋白质印迹法检测H2122SR和H2030SR中eIF4E、COPS5和ORC2的表达与亲本细胞系的比较。C具有不同MYC和orc2 mRNA表达状态的154例KRAS突变型NSCLC患者的生存图。使用对数秩检验评估统计学差异。数据来自TCGA泛癌症图谱。D用安洛替尼 (2 μ m) 加索托拉西布 (1 μ m) 处理的五种细胞系 (Calu-1,SW1573,H23,H2122SR和H2030SR) 中ORC2的表达通过western印迹检测24 °h。索托·索托罗西布,安洛·安洛替尼。EMYC过表达和ORC2敲低后H2122和H2030的细胞活力测定 (si-rna20 °m,72 °h)。结果显示为平均值 ± SEM。F-IH23的EdU/PI染色测定 (F,G) 和H2122SR (H,我) 用EV,myc-oe,si-ORC2 #1或si-ORC2 #2转染后,用安洛替尼 (2 μ m) 加索托拉西布 (1 μ m) 处理24 h (20 μ m,72 °h)。结果显示为平均值 ± SEM。使用具有Tukey多重比较检验的单向ANOVA确定统计学差异。粗体:p < 0.05。n = 3每组。组合: 安洛替尼加索托拉西布。J-MH23的细胞生长和细胞活力测定 (J,K) 和H2122SR (L,M) 用安洛替尼 (2 μ M) 加索托拉西布 (1 μ M) 处理24 h EV,myc-oe,si-ORC2 #1或si-ORC2 #2 (20 μ M,72 h)。结果显示为平均值 ± SEM。使用具有Tukey多重比较检验的单向ANOVA确定统计学差异。粗体:p < 0.05。n = 3每组。组合: 安洛替尼加索托拉西布。
对TCGA数据集 (154例KRAS突变型NSCLC患者) 的分析显示,MYC的高表达与不良的总体生存率相关 (图。
S4K) [
19]。值得注意的是,ORC2表达与MYC表达呈正相关 (图。
S4L),两个基因高共表达的患者的生存结果最差 (图。
5C)。
ORC2作为DNA复制引发剂复合物的一部分,与其他ORC亚基 (ORC1-6) 一起介导DNA复制,在致癌作用中起着至关重要的作用 [
20]。正如预期的那样,联合治疗抑制了ORC2的蛋白质表达 (图。
5D)。ORC2敲除 (图。
S5A) 从myc-oe诱导的H2122和H2030亲本细胞系的抗性中恢复sotorasib敏感性 (图。
5E)。MYC的过表达在mRNA和蛋白质水平上上调了ORC2 (图。
S4I和
S5A),ORC2敲低逆转了MYC过表达对联合治疗的保护作用,恢复了H23和H2122SR细胞的细胞周期停滞和凋亡 (图
5F-I,和
S5B-E)。同样,ORC2敲低恢复了联合治疗对MYC过表达细胞中肿瘤生长和活力的抑制作用 (图。
5J-M)。
这些发现确立了c-myc/ORC2轴作为对与KRAS-G12Ci组合的安洛替尼在克服抗性中的反应的关键介质。
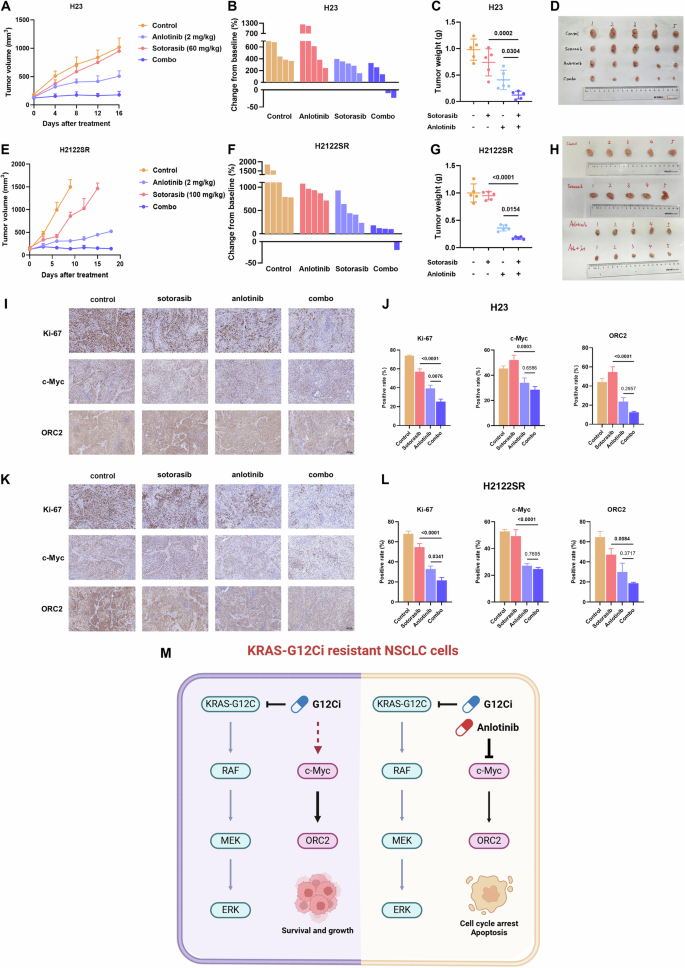
安洛替尼联合KRAS-G12Ci在体内耐药环境中发挥有效的肿瘤抑制作用为了验证我们的体外发现,我们使用H23和h2122srcdx模型评估了组合治疗的体内功效。虽然单药sotorasib显示出有限的功效,但在两种模型中,联合治疗均显示出有效的肿瘤生长抑制作用,而对体重没有显着影响 (图
6A-H和
S5F, G)。
A-H安洛替尼联合索托拉西布在H23 CDX模型中的抗肿瘤作用 (A-D) 和H2122SR CDX型号 (E-H)。结果显示为平均值 ± SEM。使用具有Tukey多重比较检验的单向ANOVA确定统计学差异。粗体:p < 0.05。n = 5每组。组合: 安洛替尼加索托拉西布。I-LKi-67、c-myc和ORC2在H23 CDX模型中的表达 (我,J) 和H2122SR CDX型号 (K,L) 通过IHC检测。结果显示为平均值 ± SEM。使用具有Tukey多重比较检验的单向ANOVA确定统计学差异。粗体:p < 0.05。n = 5每组。组合: 安洛替尼加索托拉西布。MKRAS-G12Ci耐药NSCLC细胞中由安洛替尼加索托拉西布诱导的c-myc/ORC2抑制的图形摘要。创造了智慧h Biorender.com。
肿瘤组织的免疫组织化学分析显示,联合治疗显着降低了H23和H2122SR异种移植物中Ki-67 (增殖标志物),c-myc和ORC2的表达 (图。
6I-L)。
支持这些发现,TMA分析表明ORC2和c-myc蛋白表达水平之间呈正相关。重要的是,c-myc和ORC2的高共表达与NSCLC患者更差的生存结果相关 (图。
S5H, I)。
这些体内结果以及体外结果验证了我们的机制研究结果,并证明安洛替尼与KRAS-G12Ci组合通过调节c-myc/ORC2轴有效地克服了耐药性,导致显著的肿瘤抑制 (图。
6米)。
讨论KRAS-G12C抑制剂的出现代表了NSCLC治疗的重大突破,但其临床疗效仍受到原发性和获得性耐药机制的限制 [
9,
10,
12]。我们的研究表明,安洛替尼与KRAS-G12C抑制剂的组合通过调节c-myc/ORC2信号轴有效地克服了这些耐药机制。
我们的研究结果表明,安洛替尼协同增强KRAS-G12Ci对多种耐药NSCLC细胞系的抗肿瘤作用。在初级抗性品系 (H23、Calu-1、SW1573) 和获得性抗性品系 (H2122SR、H2030SR) 中均观察到这种协同作用,表明这种组合策略的广泛适用性。增强的功效通过增加对细胞活力、长期存活和迁移的抑制来表现,表明对癌细胞行为的多方面影响。机械地,组合疗法在抗性细胞系中诱导稳健的细胞周期停滞和细胞凋亡。
转录组分析和进一步验证揭示了c-myc/ORC2轴是组合疗法效果的关键介质。C-myc及其下游靶标ORC2的下调似乎是抗肿瘤反应中的关键事件。作为DNA复制引发剂复合物的一部分,ORC2与其他ORC亚基 (ORC1-6) 一起介导DNA复制,在致癌作用中起着至关重要的作用 [
20]。有趣的是,一项针对胰腺癌的研究发现,polo样激酶1的ORC2磷酸化在压力条件下促进DNA复制,并导致吉西他滨耐药,这表明其在癌症进展中的潜在重要性 [
21]。与我们的体外结果一致,c-myc和ORC2表达之间的正相关,以及它们与NSCLC患者中较差的生存结果的关联,进一步强调了靶向该轴的临床相关性。先前的研究已经证实了MYC上调在KRAS-G12Ci抗性中的重要性以及靶向c-myc以改善KRAS-G12Ci疗效的潜力 [
11,
22]。安洛替尼已被证明在多发性骨髓瘤中直接靶向c-myc [
23],并通过靶向非小细胞肺癌中的c-met/MYC/AXL轴来逆转耐药性 [
24]。这些发现加强了我们的研究结果,即安洛替尼联合KRAS-G12Ci通过靶向c-myc/ORC2轴在KRAS-G12Ci耐药环境中的抑制作用。据报道,在多种癌症类型中,特别是NSCLC中,KRAS突变患者对安洛替尼联合治疗的临床反应 [
25,
26,
27,
28],为我们的组合策略的潜在应用提供了证据。
体内研究证实了我们的体外发现,证明了原发性耐药 (H23) 和获得性耐药 (H2122SR) 异种移植模型中的有效肿瘤抑制。肿瘤生长的显着减少,加上肿瘤组织中Ki-67,c-myc和ORC2的表达降低,为这种组合疗法的翻译潜力提供了证据。
自2018年以来,安洛替尼已被NMPA批准作为中国NSCLC患者的三线或后续治疗选择,在ALTER0303试验中证明了作为单药治疗的良好安全性。现实世界的研究进一步证实,无论是单独使用还是与PD-1抑制剂联合使用,与安洛替尼相关的不良事件在肺癌和其他恶性肿瘤中都是可控的 [
29,
30,
31,
32]。此外,多项临床试验表明,当与PD-1抑制剂或egfr-tkis联合使用时,安罗替尼作为安全有效的候选药物具有潜力,可显着延长PFS并克服转移性NSCLC患者的耐药性 [
33,
34,
35]。这些发现进一步支持了探索安洛替尼与KRAS-G12C抑制剂组合作为该患者群体的有希望的治疗策略的基本原理。
然而,应该考虑几个限制和未来的方向。虽然我们的研究集中在非小细胞肺癌,但这种组合策略对其他KRAS-G12C突变癌症的适用性需要进一步研究。此外,这种组合在患者中的长期安全性和有效性需要通过临床试验进行评估。此外,这种组合抑制c-myc的潜在机制需要澄清。未来的研究应该探索这种联合疗法的潜在耐药机制和克服它们的策略。
总之,这项研究为临床评估安洛替尼与KRAS-G12C抑制剂联合治疗对KRAS-G12Ci单药治疗原发性或获得性耐药的NSCLC患者提供了有力的依据。C-myc/ORC2轴作为反应的关键介质的阐明为KRAS-G12Ci抗性的分子基础提供了新的见解,并提出了新的治疗靶标。这种组合策略有望改善KRAS-G12C突变NSCLC患者的预后,有可能将精准医学的益处扩展到更广泛的患者群体。
数据可用性Rna-seq数据已经保存到cncb-ngdc数据库 (HRA010830)。根据通讯作者的合理要求,可以获得本研究的数据。
 妈妈与脑膜转奋战4年多,如今也成功
作者:李妮妮
2020年12月1号,妈妈在确诊晚期肺癌4年零10月,经历了化疗、一代靶向易
妈妈与脑膜转奋战4年多,如今也成功
作者:李妮妮
2020年12月1号,妈妈在确诊晚期肺癌4年零10月,经历了化疗、一代靶向易
 三代伏美换一代特罗凯获益率问题
家父25年2月因肩膀疼痛确诊肺腺癌晚期4B,双肺转,多发骨转,脑转(非典型),
三代伏美换一代特罗凯获益率问题
家父25年2月因肩膀疼痛确诊肺腺癌晚期4B,双肺转,多发骨转,脑转(非典型),
 右肺上叶切除,术前术后对比
今年2月21日手术前发现占位40×31,术后辅助治疗发现团块影82×75?谁能帮我看看两份
右肺上叶切除,术前术后对比
今年2月21日手术前发现占位40×31,术后辅助治疗发现团块影82×75?谁能帮我看看两份
 5年奥希还是复发了,伴有恶性胸水该
家母2020年直接开始服用奥西替尼,效果一直很好,最近两个月出现胸水以及锁骨淋巴肿大
5年奥希还是复发了,伴有恶性胸水该
家母2020年直接开始服用奥西替尼,效果一直很好,最近两个月出现胸水以及锁骨淋巴肿大
 父亲奥西+卡马耐药后应该如何选择?
2021年9月确诊肺腺癌,基因检测结果EGFR19缺失,服用奥希替尼。2024年3月CT影像检查出
父亲奥西+卡马耐药后应该如何选择?
2021年9月确诊肺腺癌,基因检测结果EGFR19缺失,服用奥希替尼。2024年3月CT影像检查出



 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡


